Caryodaphnopsis Airy Shaw
Revisión
van der Werff, H. & H. G. Richter. 1985. Caryodaphnopsis Airy-Shaw (Lauraceae), a genus new to the Neotropics. Systematic Botany 10: 166–173.
Descripción
El género Caryodaphnopsis se caracteriza dentro de la familia Lauraceae por sus hojas
opuestas, usualmente trinervadas (caracteres excepcionales en la familia), raramente
peninervias; inflorescencias tirsopaniculadas, flores trímeras, bisexuales (Kubitzki, 1994) y seis tépalos marcadamente desiguales, siendo los externos mucho más pequeños que los
internos (van der Werff & Richter, 1996).
Adicionalmente, la mayoría de las especies de Caryodaphnopsis poseen anteras de 4 locelos (en 2 especies anteras de 2 locelos), sacos polínicos arreglados en arco; nueve
estambres fértiles (raramente 6), el tercer verticilo con glándulas, filamentos más largos o
más cortos que las anteras, los estaminodios del cuarto verticilo conspicuos, de cordados
a sagitados. Además presenta un receptáculo muy pequeño y aplanado, pedicelo no ensanchado y tépalos deciduos o subpersistentes (Kubitzki, 1994).
Algunas de las características de este género como anteras de 4 lóculos, nueve estambres
fértiles y hojas trinervadas, distinguen a Caryodaphnopsis entre la familia Lauraceae, ya
que la mayoría de géneros poseen anteras de 2 lóculos, seis estambres fértiles y hojas
peninervias (van der Werff & Richter, 1985). Adicionalmente, Caryodaphnopsis posee
una estructura anatómica de madera única dentro de la familia Lauraceae (van der Werff & Richter, 1985). Sin embargo, se ha enfatizado en las hojas opuestas, los tépalos
desiguales y la estructura única de la madera (van der Werff & Dao, 1999), como
caracteres diagnósticos a nivel genérico, que determinan la monofilia de Caryodaphnopsis dentro de la familia Lauraceae (van der Werff & Richter, 1996).
Historia taxonómica
Este género, reportado originalmente para Asia, fue descrito por Airy Shaw en 1940,
quien creó el nombre del género debido a similitudes entre las hojas de éste con las hojas
de algunas especies de Cryptocarya (=Caryodaphne) (Kostermans, 1974). En el caso de
las especies C. tonkinensis y C. baviensis fueron descritas originalmente por Lecomte
(1913) bajo el género Notaphoebe, hasta 1940 cuando A. Shaw las incluyó dentro de
Caryodaphnopsis, y que Kostermans las ratificó dentro de éste género en 1964, después de haber pasado por otros géneros. Posteriormente fueron añadidas al género Caryodaphnopsis las especies C. latifolia y C. laotica por W.F Wang y A. Shaw en 1957 y 1960 respectivamente (Kostermans, 1974).
En 1952 el género Caryodaphnopsis fue ubicado en sinonimia bajo el género Persea, junto con los géneros Alseodaphne y Notaphoebe, debido a la relación cercana que existe
entre estos. Sin embargo, las diferencias que se encuentran entre estos géneros fueron lo
suficientemente fuertes para que Caryodaphnopsis fuera delimitado como un género separado, en 1974 por Kostermans. Este último incluyó en su monografía del género siete especies, cuyo rango de distribución iba de Yunnan a Borneo y Filipinas
(Kostermans, 1974).
Hasta 1985 este género sólo estaba reportado para Asia Tropical, pero van der Werff y Richter, transfirieron dos especies Suramericanas de Persea al género Caryodaphnopsis
(Zamora et al, 1988). Esta transferencia entre géneros se dio debido a evidencias que
soportaban la presencia de las especies Persea theobromifolia Gentry y Persea inaequalis
A.C Smith en el género Caryodaphnopsis, ya que éstas diferían de otras especies del
género Persea en cuanto a morfología (foliar y floral) y estructura de la madera (Van der
Werff & Richter, 1985), siendo éstas características más congruentes con el género Caryodaphnopsis.
Distribución
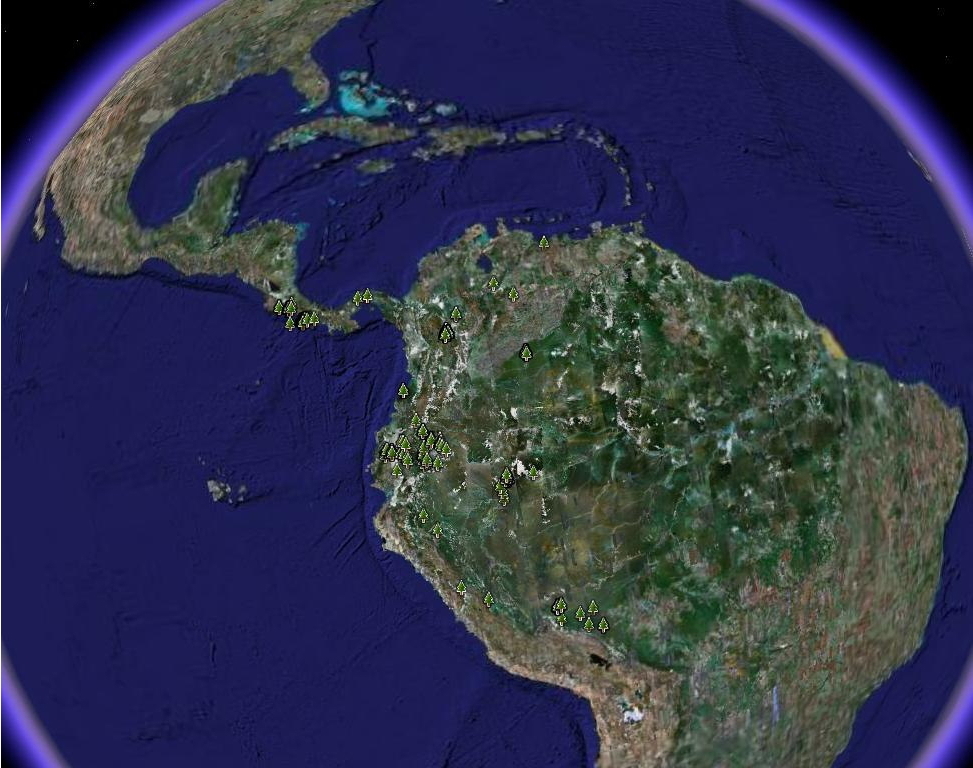
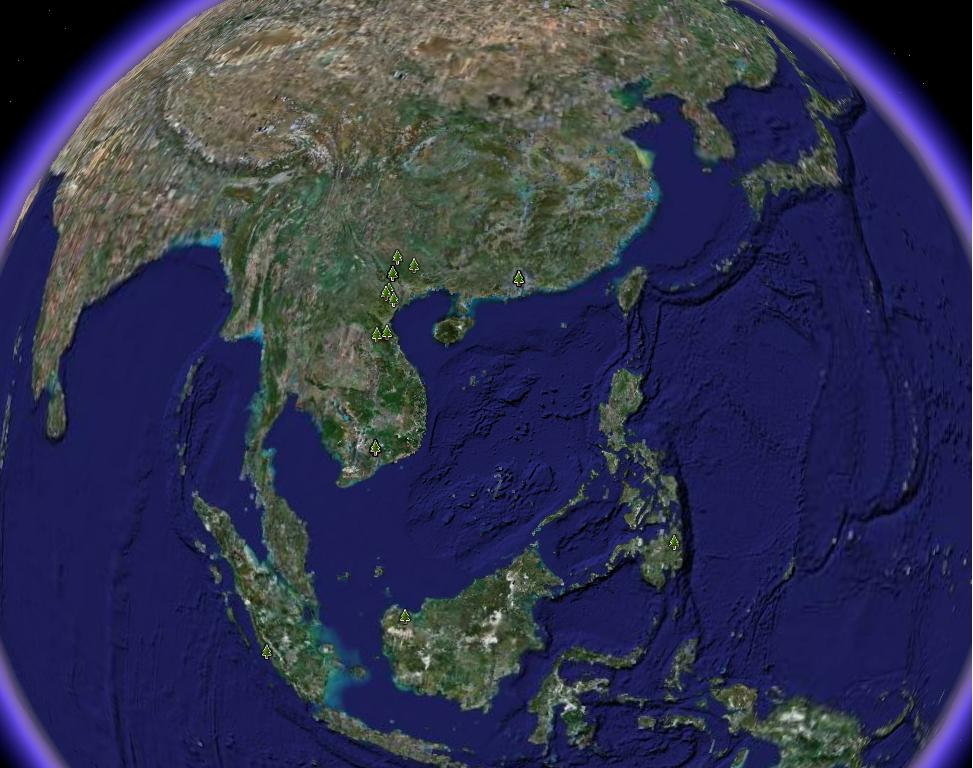
El género Caryodaphnopsis, cuenta actualmente con 15 especies. Es un género tropical
anfi-Pacífico (van der Werff & Richter, 1985), que presenta una marcada disyunción
geográfica. Esta ocurre entre el Sudeste Asiático (Sur de China, Vietnam, Laos,
Cambodia, Indonesia y Filipinas) y América tropical (Brasil, Perú, Ecuador, Colombia,
Venezuela y Costa Rica) (Aymard & Romero-González, MS)
Gracias al descubrimiento de una especie de Trigonobalanus (actualmente Colombobalanus) , perteneciente a la familia Fagaceae en Colombia, género
anteriormente reportado exclusivamente para Asia y que presentaba un claro ejemplo de
disyunción anfi-pacífica, Van der Hammen y Cleef (1983) concluyeron que los taxa que presentan este tipo de distribución, en el pasado tenían una distribución mucho más
amplia en el hemisferio norte durante el cálido Terciario y posteriormente fueron
forzados a migrar hacia el sur debido al enframiento generalizado durante el Terciario
tardío y los periodos glaciales.
Posición sistemática
Estudios sobre la posición filogenética del género Caryodaphnopsis dentro de la familia Lauraceae sugieren tres diferentes escenarios: 1. Según Chanderbali et al. (2001), Caryodaphnopsis es el género más enigmático de la familia Lauraceae, junto con Neocinnamomum y Cassytha, ya que con base en análisis combinados de los genes trnL-trnF y psbA-trnH, estos tres géneros constituyen un clado, cercanamente relacionado al clado Cryptocaryeae y más aún al clado Chlorocardium-Mezilaurus y al clado Perseeae-Laureae (Figura 2a). 2. Con un segundo análisis combinado de los genes trnL-trnF, psbA-trnH, trnT-trnL, rpl16 y 26S realizado en el mismo estudio de Chanderbali et al. (2001), estos géneros ya no forman un clado, pero su posición dentro del árbol de la familia se mantiene, con un alto soporte de bootstrap (Figura 2b). Sin embargo las relaciones que puedan existir entre estos géneros aún no son bien entendidas debido a que morfológicamente existen diferencias muy pronunciadas. Con Neocinnamomum excluido del análisis, Cassytha y Caryodaphnopsis constituyen un clado. Siendo Cassytha el único género de plantas parásito de la familia, lo único que tiene en común con Caryodaphnopsis es el verticilo externo de tépalos extremadamente reducido, lo cual no puede ser considerado como una característica determinante, ya que se presenta constantemente dentro de la familia. Hasta el momento, no se han encontrado otros caracteres que soporten la relación entre estos dos géneros (Chanderbali et al. 2001). Con respecto a Neocinnamomum, éste género comparte una morfología similar con el género Caryodaphnopsis, ambos teniendo venación trinervada y anteras de 4 locelos dispuestos en arco (raramente anteras con 2 locelos en Caryodaphnopsis). Sin embargo, existen características que no comparten, como por ejemplo la disposición de las hojas, perianto y cúpula: Caryodaphnopsis presenta hojas opuestas, posee un perianto con tépalos fuertemente desiguales y carece de cúpula, mientras que Neocinnamomum presenta hojas alternas, un perianto con tépalos casi iguales, y presenta una cúpula con tépalos persistentes (Chanderbali et al. 2001). Por lo tanto, las relaciones encontradas dentro del clado Caryodaphnopsis-Neocinnamomum-Cassytha, no presentan un fuerte soporte morfológico, contrariamente a lo que ocurre con la ubicación basal de este clado dentro del árbol de la familia, ya que su posición entre el clado Cryptocaryeae y el clado Chlorocardium-Mezilaurus, se ve fuertemente soportada por afinidades anatómicas que existen entre Neocinnamomum y Caryodaphnopsis, con Chlorocardium y Cryptocaryeae, respectivamente (Richter, 1981). 3. Al remitirse a estudios previos como el de Rohwer del 2000, en el que se analizan evidencias moleculares basadas en secuencias obtenidas del gen matK (cloroplasto), se puede observar una posición sugerida de Caryodaphnopsis entre el clado Cryptocaryeae y los clados Chlorocardium-Mezilaurus- Williamodendron y Perseeae-Laureae (Figura 2c). Los resultados encontrados en este estudio se complementan con los encontrados por Chanderbali et al. (2001), con el género Williamodendron como un componente nuevo del clado. Sin embargo difieren en la posición del género Cassytha, que en el estudio de Rohwer (2000) presenta una posición basal en el árbol de Lauraceae, un poco alejada de Caryodaphnopsis. Con respecto a Neocinnamomum, género que se agrupaba con Caryodaphnopsis en el estudio de Chanderbali et al. (2001), no fue incluído en los análisis de Rohwer (2000), sin embargo este último autor sugiere que morfológicamente estos dos géneros están cercanamente relacionados. Según los resultados obtenidos por Rohwer (2000), el grupo hermano de Caryodaphnopsis presenta un fuerte soporte de bootstrap. Los anteriores acercamientos a la posición sistemática de Caryodaphnopsis, son las 3 hipótesis iniciales en las que se basa el presente estudio, que tiene como propósito la determinación de la posición de este enigmático género dentro de la filogenia de la familia Lauraceae.
Discusión
La presencia de Caryodaphnopsis como un grupo monofilético es un elemento común a
las filogenias generadas con los diferentes genes. La conformación del género como un
clado bien soportado, se apoya con los caracteres morfológicos usados para la
clasificación de Caryodaphnopsis a nivel genérico. Una buena clasificación demanda que
los caracteres usados sean confiables, lo que significa que no hay o hay pocas
excepciones en las condiciones características de un taxón. El análisis de los caracteres
usados para la clasificación permite estimar qué tan bien fundamentado se encuentra un
taxón (van der Werff & Richter, 1996). Caracteres que son muy variables dentro de los
géneros de Lauraceae sólo pueden ser utilizados en géneros que son definidos por otros
caracteres (van der Werff & Richter, 1996). Tal es el caso de Caryodaphnopsis, que a
nivel genérico se define por su disposición opuesta de las hojas, sus tépalos desiguales y
su estructura única de la madera, y que a pesar de presentar variabilidad en características
como número de estambres y número de locelos en los estambres, puede considerarse
como un género monofilético (van der Werff & Richter, 1996). El caso de
Caryodaphnopsis es un claro ejemplo de monofilia soportada por evidencia molecular y
morfológica, lo que quiere decir que con total seguridad es un género muy bien
fundamentado.
En los análisis realizados con los marcadores psbA-trnH, matK e ITS, donde el muestreo
de Caryodaphnopsis fue mayor a dos especies, se pudo observar la diferenciación del
grupo americano con respecto al grupo asiático.
Contrario a psbA-trnH, la variabilidad ofrecida por el gen ITS fue muy
valiosa para resolver las relaciones filogenéticas de las especies de Caryodaphnopsis.
Teniendo en cuenta que ITS parece ser un marcador efectivo como diagnóstico molecular
para identificación a nivel específico, puede ser propuesto para la inclusión en el actual
proyecto de Códigos de Barra de la Vida.
A pesar de ser un análisis preliminar debido a la falta de muestreo de algunas especies, el
marcador ITS resolvió relaciones dentro del grupo de las especies americanas de
Caryodaphnopsis, siendo consistentes con evidencia morfológica reportada por van der
Werff en 1988. Este autor sostiene que existe una tendencia de las especies neotropicales
a agregarse en dos grupos definidos por caracteres morfológicos como nerviación de las
hojas y forma del fruto.
Estas características determinan la conformación del clado C. burgeri-C.fosteri que
presenta sinapomorfias como hojas trinervadas y frutos pequeños y globosos. En este
clado se incluyeron dos especies no determinadas: Caryodaphnopsis sp. nov 2 y
Caryodaphnopsis aff. fosteri.
En el caso de C. sp. nov2 se trata de un nuevo registro para Río Claro, Antioquia
(Colombia) y con base en descripciones morfológicas se presumía que era el primer
registro para el país de Caryodaphnopsis burgeri (Cogollo, com. pers). Según los
resultados obtenidos con evidencia molecular del marcador ITS, se sugiere que C.sp.nov 2 se trata en efecto de C. burgeri. Acerca de C. aff. fosteri se evidencia en los análisis una
cercanía con C. fosteri, por lo tanto esto apunta a que estos dos especimenes pertenecen a
la misma especie.
Adicionalmente, la conformación del clado C. cogolloi-C. theobromifolia con base en la
evidencia molecular, se apoya con las observaciones de van der Werff (1988), el cual
sostiene que este grupo tiene como sinapomorfias las hojas peninervias y los frutos con
forma de aguacate (obovoide). Debido a problemas de amplificación y secuenciación no
fue posible incluir el tercer miembro de este grupo, C. inaequalis, lo cual hubiera sido
muy interesante para seguir evaluando la efectividad de ITS y ver si se mantenía la
congruencia entre la evidencia morfológica con respecto a la evidencia molecular.
Con respecto de la posición de Caryodaphnopsis dentro de la familia Lauraceae al
evaluar las hipótesis iniciales y las filogenias obtenidas mediante los diferentes genes
muestra cierta tendencia, no obstante, no se puede inferir nada determinante.
La relación cercana entre Caryodaphnopsis y los clados Chlorocardium-Mezilaurus y
Perseeae-Laureae, es un elemento planteado en las hipótesis evaluadas inicialmente, y es
una situación que se presenta en los análisis realizados con los genes psbA-trnH, matK,
rpl16, 5.8S y trnT-trnL. No obstante existen factores como bajos soportes de bootstrap y
creación de politomías por colapso de ramas, que le añaden incertidumbre a las relaciones
filogenéticas obtenidas con los diferentes genes evaluados.
La posición de Caryodaphnopsis con respecto al clado Cryptocaryeae fue variable en los
análisis ya que Cassytha y Neocinnamomum eran determinantes para esto y se pudo ver
que a través de todos los análisis la posición de estos géneros fue muy variable.
Al examinar los genes utilizados en los análisis, se pudo determinar que matk fue el único
gen que presentó soportes de bootstrap altos y en donde no se presentaron politomías.
Además ofreció una filogenia congruente con la segunda hipótesis inicial, lo que sugiere
que este marcador es efectivo para la construcción de filogenias a nivel de clados
profundos en las Lauraceae.
